یک کِرم میکروسکوپی ممکن است به چگونگی درک ما از گرانش کمک کند
درحالیکه انسان، برای حفظ تعادل و جهتیابی، به گرانش وابسته است، مکانیسمی که ما، بهواسطهٔ آن، نیروی بنیادی گرانش را حس میکنیم تا حد زیادی ناشناخته مانده است؛ حتی عجیبتر از آن، ارگانیسم مدل کرم سی. الگانس (یک کرم میکروسکوپی) نیز میتواند جهت گرانش را حس کند؛ گرچه هیچ دلیل زیستمحیطی برای این امر شناخته نشده است.
گروهی از محققان مهندسی پن، با مدیریت هایم بائو استاد مهندسی مکانیک و مکانیک کاربردی و دیوید رایزن دانشیار نورولوژی در مدرسه پزشکی پرلمن، برای پردهبرداشتن از این راز و رسیدن به بنیادیترین احساس ما از گرانش، سلسله آزمایشهایی را بر روی این ارگانیسم مدل انجام دادند. همچنین، آلکس چن دانشجوی دکترا و هونگتانگ کو دانشجوی کارشناسیارشد و اُسوالد چوانگ همکار پسادکترا در این پژوهش مشارکت کردند.
کرم سی. الگانس، با وجود سادگی فیزیولوژی بسیارش، بیش از نیمی از ژنهایش با انسان مشترک است، که این امر سبب میشود مطالعات ژنتیکی منجر به شناخت عمیق ژنهای مسئول در رفتارهای مشابه در انسان شوند. اما فکر اولیهٔ مطالعهٔ ژنتیک گراویتاکسی (gravitaxis)، یا توانایی حرکت در واکنش به نیروی گرانش، کاملاً اتفاقی بود. این به معنای آن است که محققان از رفتار این کرمها آگاه نبودند.
بائو میگوید: «ما در حال مطالعه دربارهٔ موضوعی دیگر بودیم؛ یعنی هیدرودینامیک کرم سی. الگانس. بدینمنظور، آنها را در آب رها کردیم. وقتی مشاهده کردیم که کرمها همگی در حال رفتن به سمت کف ظرف آزمایش هستند، این پرسش برایمان ایجاد شد که آیا آنها در حال واکنش به گرانش هستند یا به دلیل سنگینی، منفعلانه در حال غرقشدن هستند.»
این گروه کار خود را با تحقیق دربارهٔ گذرگاه مولکولی مسئول برای گراویتاکسی، آغاز کرد که این تحقیق در مجلهٔ BMC Biology به چاپ رسید.
بائو میگوید: «کرم سی. الگانس، بهطور ویژه، قادر به انجام دستکاریهای ژنتیکی است؛ بهطوریکه ما میتوانیم ژنها را فعال یا غیرفعال کنیم. این موضوع به ما در مورد عملکرد کرمها اطلاعاتی میدهد که در نتیجهٔ آن میتوان این دانش را برای ژنهای انسان نیز به کار برد، چرا که این مکانیسمهای مولکولی در قلمرو حیوانی دوام آوردهاند.»
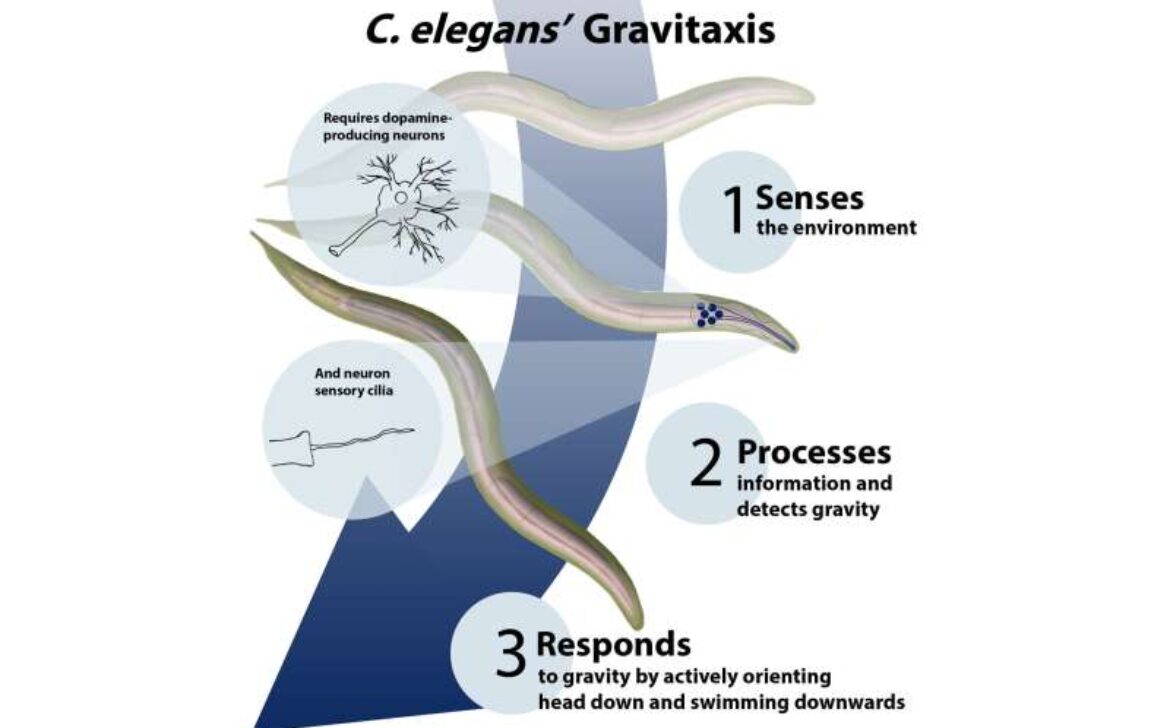
با محلولهای آبی، که هم پُرچگالتر و هم کمچگالتر از کرم بودند، آزمایشهایی انجام شد تا توانایی آن در شناسایی و شناخت گرانش بررسی شود.
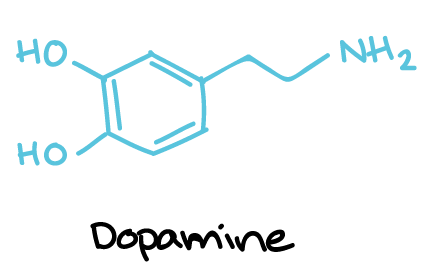
بائو میگوید: «در عرض چند ثانیه، کرمها به سمت پایین چرخ خوردند و به سمت کشش گرانش شناور شدند. ما مشاهده کردیم که واکنش کرمها فعالانه و خودخواسته بود و نه غرقشدنی منفعلانه. ما، سپس، دیگر متغیرها، از جمله وجود مژکهای حسی و نورونهای دوپامینرژیک سالم را بررسی کردیم.»
برای شناسایی سازوکار (مکانیسم) واکنش کرمها به گرانش، محققان در ابتدا عملکرد مژکهای حسی را مختل کردند -که ساختارهای سلولی مسئول در عملکردهای حسی مثل مزه، بو و دما بودند. کرمها، بدون این مژکها دیگر به سمت پایین شناور نشدند و با گرانش همراستا نگردیدند. سپس، محققان دریافتند که کرمهای فاقد انتقالدهندهٔ نورونی دوپامین، یا گیرندههایی که به دوپامین واکنش میدهند، فاقد توانایی تشخیص گرانش نیز هستند.
بائو میگوید: «ما متغیرهای ژنتیکی را ــ هم بهمنظور تعیین متغیرهای مسئول و کاربردی و هم بهمنظور کاستن متغیرهایی که نتایج آزمایش را از مسیرش منحرف میکنند ــ بررسی میکنیم. ما وقتی که میخواهیم ژنها را در جهشهای بسیارِ کرم سی. الگانس فعال و غیرفعال کنیم، در ابتدا با فرضیهٔ عملکرد ژن ــ که از یافتههایمان در رابطه با ژنی خاص داریم ــ شروع میکنیم. ما میدانیم که دوپامین یک انتقالدهندهٔ نورونی رایج است که بسیاری از کارکردهای بدن را کنترل میکند؛ بنابراین، وقتی ما آن را در جهشها غیرفعال کردیم، توانایی شناسایی گرانش از بین رفت.
او همچنین اضافه کرد: «جالب است که وقتی کرمها، در مرحلهٔ رشد کرمینهای، در معرض مکملهای دوپامین و سپس در محلول قرار گرفتند، تا حدی توانایی حسی آنها بازگشت، که این نشان میدهد ترمیم دارویی میتواند ممکن باشد.»
ارتباط بین دوپامین و احساس گرانش میتواند منظری برای برنامههای کاربردی در سلامت انسان فراهم کند.
بائو میگوید: «تحقیق ما در مرحلهای ابتدایی قرار دارد و هنوز تا برنامههای کاربردی مربوط به سلامت انسان، مسیری طولانی در پیش است. هرچند، قطعاً، برنامههای کاربردیِ حاصل از این مطالعه ــ که فراتر از کنجکاوی علمی باشد ــ وجود دارد. یافتههای ما، تحقیقهای آینده دربارهٔ احساس گرانش در انسان را تسهیل میکند؛ مخصوصاً این یافتهها برای اشخاص پیرتر ــ که در معرض افتادن هستند ــ مفید است.
بهعنوانمثال، بیماران مبتلا به پارکینسون ــ که نورونهای دوپامین معیوب دارند ــ مکرراً در معرض افتادن هستند، که این احتمالاً ناشی از عدم احساس گرانش است. بنابراین، دانستنِ اینکه چگونه دوپامین و احساس گرانش در کرمها با یکدیگر در تعامل قرار میگیرند، ممکن است با هدف یافتن کاربرد آن برای انسان باشد.
ترجمه: علی سیدآبادی
منبع:
https://phys.org/news/2021-09-microscopic-worm-gravity.html